The prevalence of obesity and overweight are increasing and is a worldwide health epidemic.1 Obesity is a complex disorder, which results from an imbalance between energy intake and expenditure, in which excessive body fat has accumulated to the extent that it may have a significant influence on morbidity and mortality.2 Twin and family studies have demonstrated that genetic factors may also contribute to levels of physical activity and eating behaviors, which ultimately affect obesity. The estimates of the heritability of body mass index (BMI) is significantly high (30 to 70%).2,3 In recent decades, there has been an impressive propagation in our knowledge base regarding obesity. It was found that the genetic components of obesity are key contributors to individual risk.4 Obesity is a global issue with no current effective treatment.5 Changes in diet cause the increasing prevalence of non-communicable diseases in developing countries, the Middle East, and North Africa so that 77.9% of chronic diseases are related to these countries.6 Based on a systematic analysis of studies in 2008 on the epidemiology of obesity in 199 countries, 1.46 billion adults were overweight and 502 million were obese.7 According to an Iranian health study conducted in 2005, the prevalence of overweight and obesity was 42.8% in men and 57% in women. Sex, age, socioeconomic factors, physical activity, smoking status, number of children, and urbanization are the main unrelated factors associated with adult obesity in Iran.8 Obesity is gaining acceptance as a very serious primary health burden, which impairs quality of life because of its associated complications, such as diabetes, cardiovascular disease, cancer, asthma, hepatic impairment, renal dysfunction, sleep disorder, and infertility.9
Genome-wide association studies are used as prescreening tools for the detection of genetic variants associated with obesity and other related diseases.10,11 It has been suggested that obesity-related genes may be involved in energy intake and expenditure. Two obesity-associated candidate genes are the fat mass and obesity-associated (FTO) gene and the melanocortin-4 receptor (MC4R) gene.2,12,13 FTO is one of the members of the AlkB family of non-heme Fe (II) and 2-oxoglutarate dependent dioxygenases, which are involved in the repair of DNA alkylation damage.14 Human or mice FTO protein have been shown to demethylate 3-methylthymine (3-metT) in single-stranded oligonucleotides (ssDNA) and 3-methyluracil (3-meU) in single-stranded RNA in vitro.15 In vivo studies have confirmed the role of FTO in energy homeostasis, as FTO knockout mice exhibit intensive decreased weight, the delay in growth, destruction of white adipose tissue, and eventually death.16–18 In contrast, FTO overexpression in mice leads to increased food intake and fat mass.19 Nevertheless the underlying link between the putative demethylase function of FTO and/or energy homeostasis remains unknown.14 In humans, the MC4R gene is located on chromosome 18, and similar to the FTO gene, plays a regulatory role in overweight status.20 MC4R is a component of the leptin system, which is expressed in the brain and is part of the melanocortin signaling pathway and is known to play an important role in control of food intake and metabolic rate.12 Various studies indicated that the MC4R rs17782313 variant is related to high energy intake, dietary fat, weight change, and risk of obesity-related diseases.21–25
The aim of our study was to investigate the association between FTO (rs9939609) and MC4R (rs17782313) polymorphisms as possible genetic factors in individual susceptibility to obesity.
Methods
A total of 213 Iranian volunteers were enrolled in the study including 130 obese and 83 healthy individuals. The study groups were selected regardless of gender, physical activity levels, and family history of obesity. All cases were aged over 20 years old. The study groups were classified into three groups according to their BMI: normal/healthy weight (18.5 – < 25), overweight (25 – < 30), and obese (> 30).
All patients gave their informed consent, and all procedures were approved by the Ethics Committee at Nour Danesh Institute of Higher Education, Isfahan, Iran. Briefly, 3 mL peripheral blood was collected and transferred to the lab in a sterile falcon tube.
The polymerase chain reaction (PCR) primers used were designed with Oligo 7 Software after the alignment of available GenBank sequences. The primers used are given in Table 1. Total genomic DNA was extracted from peripheral blood using the GeNet Bio extraction kit (GeNet Bio, Makrozhen, Korea) according to the manufacturer’s instructions. The concentration and purity of the DNA extracted from each sample were determined followed by OD 260/280 spectrophotometry (NanoDrop, DeNovix Inc, Wilmington, DE, USA) as well as DNA qualitative assessment on 1.5% agarose gel.
Table 1: Polymerase chain reaction primers sequence.
|
FT0 rs9939609 inner |
CCTTGCGACTGCTGTGAATATA |
CAGAGACTATCCAAGTGCATCTCA |
|
FT0 rs9939609 outer |
GCTGCTATGGTTCTACAGTTCCA |
TGTTCAAGTCACACTCAGCCTC |
|
MC4R rs17782313 inner |
GAAGTTTAAAGCAGGAGAGATTGTATACC |
GCTTTTCTTGTCATTTCCAGCA |
FTO: fat mass and obesity-associated; MC4R: melanocortin-4 receptor.
Table 2: Polymerase chain reaction temperature protocols.
|
FTO |
95 oC - 5 min |
95o C - 30 sec |
58 oC - 30 sec |
72 oC - 30 sec |
72 oC - 5 min |
FTO: fat mass and obesity-associated; MC4R: melanocortin-4 receptor.
Tetra-primer amplification refractory mutation system (ARMS)-PCR was performed using Biometra GmbH System (Biometra GmbH, Kat#846-X070-141, Makrozhen, Korea). The amplification was performed in a 25 µL reaction mixture containing 1 µL of DNA, 0.7 μL of each outer FTO primer, 1 µL of each inner FTO primer, 3 µL of each inner MC4R primer, 1 µL of each inner MC4R, 0.5 µL of dNTPs, 2.5 µL of buffer (10X), 1 µL MgCl2 (1Mm), and 0.2 µL Taq polymerase. Both PCR assays were run under the optimized conditions [Table 2].
PCR products were assessed by electrophoresis on a 1.5% agarose gel stained with SYBR® Safe DNA gel stain (Invitrogen). Also PCR products sequencing were performed on an ABI PRISM® 3100 Genetic Analyzer machine (Applied Biosystems, Thermo Fisher Scientific, and Waltham, MA, USA). The variants analysis were evaluated using Finch TV Software (PerkinElmer Inc., Waltham, MA, USA).
Statistical analyses were performed using SPSS (SPSS Inc. Released 2007. SPSS for Windows, Version 16.0. Chicago, SPSS Inc). Descriptive analyses were expressed as mean±standard deviation. The chi-squared tests and odds ratios (OR) was used to compare the proportions of the groups. Comparisons were considered statistically significant if p < 0.050.
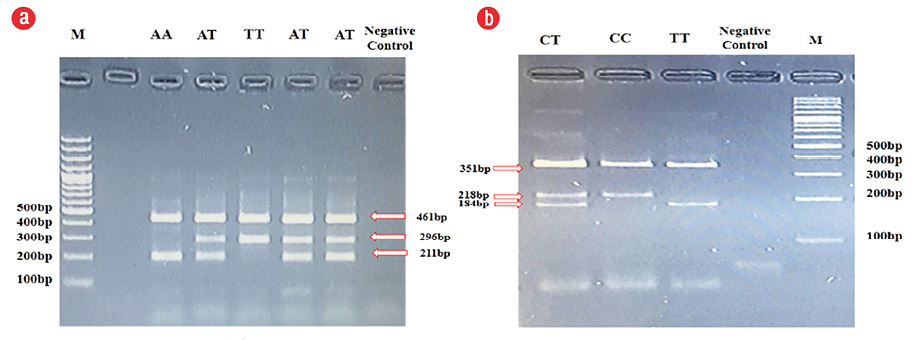
Figure 1: Electrophoresis of polymerase chain reaction (PCR) products on agarose gel 1.5% using the Tetra ARMS-PCR for the (a) fat mass and obesity-associated and (b) melanocortin-4 receptor genes. M: DNA ladder (100 bp).
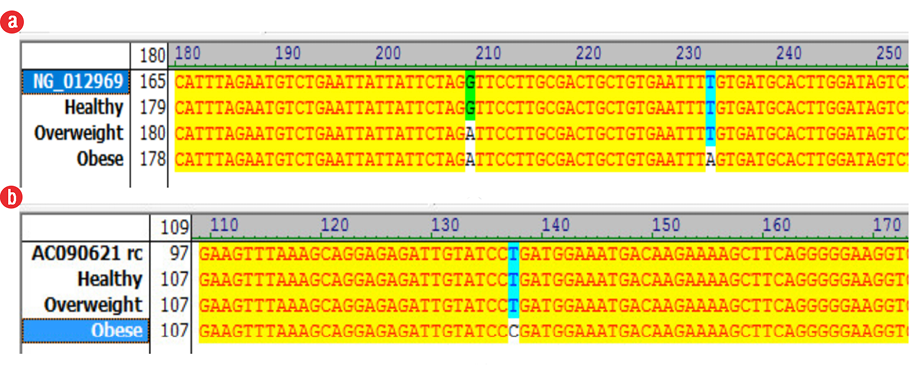
Figure 2: Nucleotide alignment of fat mass and obesity-associated (FTO) and melanocortin-4 receptor (MC4R) genes, matching of the above sequences with the target sequence (Accession no.NG_012969 and AC090621 rc) was confirmed by Vector NTI software. (a) FTO and (b) MC4R genes.
Results
Two hundred and thirteen adults were genotyped. Fifteen (23%) were homozygous for the obesity risk allele (A-A) and for the FTO SNP rs9939609, 21 (32%) were heterozygous (A-T), and 29 (45%) were wild type (T-T).
In patients with the FTO rs9939609 variant, the proportion of homozygous A-A and T-T carriers was significant (p < 0.005). Those with the FTO risk allele (A-A) had significantly higher odds of being overweight (OR = 4.269, p < 0.005, 95% CI: 2.13–8.52) [Table 3] or obese (OR = 6.927, p < 0.005, 95% CI: 3.48–13.78) [Table 4] than healthy weight/control group individuals. Moreover, no significant association between MC4R rs17782313 and obesity were observed when compared to individuals with a healthy BMI. Compared to healthy weight patients, those with MC4R risk allele (C-C) had higher odds of obesity than individuals with the T alleles (T-T and T-C) (OR = 1.889, p = 0.077, 95% CI: 0.92–3.84) [Table 3]. Analysis of the Tetra ARMS-PCR products on agarose gel showed 296 bp and 211 bp bands lane for rs9939609 and rs17782313, respectively [Figure 1]. The results obtained from sequencing confirmed the above results. To detect mutations of the FTO and MC4R genes, introns sequenced and analyzed. Sequencing results of intron 1 of the FTO gene revealed two SNP at the positions of 207 (G to A) and 231 (T to A). Also, an SNP was found at position 135 (T to C) of the MC4R gene [Figure 2].
Table 3: Summary of detected nucleotide variations and comparison overweight patients (BMI 25 – < 30) with healthy weight/control group (BMI 18.5 – < 25)
|
A-A (FTO) |
6 |
10 |
< 0.005 |
4.269 |
|
T-T (FTO) |
28 |
43 |
< 0.005 |
0.171 |
|
A-T (FTO) |
31 |
47 |
ns |
1.224 |
|
T-T (MC4R) |
14 |
22 |
ns |
0.493 |
|
C-C (MC4R) |
21 |
32 |
0.077 |
1.889 |
A p-value < 0.005 was considered statistically significant; ns: non-significant. BMI: body mass index; FTO: fat mass and obesity-associated;
MC4R: melanocortin-4 receptor.
Table 4: Summary of detected nucleotide variations and comparison obese patients (BMI > 30) with healthy weight/control group (BMI 18.5 – < 25)
|
A-A (FTO) |
15 |
23 |
< 0.005 |
6.927 |
|
T-T (FTO) |
29 |
45 |
< 0.005 |
0.244 |
|
A-T (FTO) |
21 |
32 |
ns |
0.624 |
|
T-T (MC4R) |
27 |
42 |
0.064 |
0.587 |
|
C-C (MC4R) |
17 |
26 |
ns |
1.079 |
BMI: body mass index; FTO: fat mass and obesity-associated; MC4R: melanocortin-4 receptor; ns: non-significant.
Discussion
We analyzed the SNPs rs9939609 of the FTO gene and rs17782313 of the MC4R gene in a group of obese and normal-weight Iranian patients. Our study showed that FTO genetic polymorphism increase the risk of obesity in our population. However, it is important also to consider that lifestyle factors may modulate the obesity risk associated to FTO. A recent study reported that FTO rs9939609 SNP was significantly associated with BMI (p = 0.01), weight (p = 0.03), and waist circumference (p = 0.04).26 It has been suggested that FTO gene variants have a significant association with obesity; however, the mechanisms behind this association is not yet clear. Additionally, the obesity gene FTO may influence the methylation level of other genes. It has been suggested that the obesity gene FTO is correlated with methylation changes in multiple sites, where the effect of the FTO risk allele (rs9939609) can be mediated, at least in part, via epigenetic modifications.27 The rs9939609 SNP located in the first intron of the FTO gene is of interest in the field of obesity.28 Homozygous loss-of-function mutations in the FTO causes severe growth retardation and multiple abnormalities whereas the loss of one functional copy of this gene is compatible with both obese and lean phenotypes. Leanness, postnatal growth retardation, and a higher metabolic rate were shown in FTO knockout mice, and in mice with a missense mutation in exon 6.29,30 It has been determined that both genetic and non-genetic factors contribute to the development of obesity and the risk of metabolic syndrome.31 Our study indicated that FTO rs9939609 variant was significantly associated with BMI. Also, the genotype distribution of AA-homozygotes was significantly higher in people with obesity compared to normal-weight individuals, with an increased OR (OR = 6.927, p < 0.005). Like our results, another study showed that A-A genotype carriers have a two-times higher risk for obesity compared with A-T and T-T genotype carriers.32
The influence of FTO rs9939609 and MC4R rs17782313 polymorphisms on obesity was previously investigated. The authors of the study found a significant connection between the FTO risk genotypes (AA + AT) and BMI (p = 0.03), and the MC4R risk genotypes (CC + CT) were associated with a greater BMI (p = 0.03).33
In agreement with our study, the authors of a different study showed that the FTO rs9939609 A-A genotype was significantly higher in the obese population compared to normal-weight subjects.34 Furthermore, the authors did not observe a significant association between MC4R polymorphisms and BMI.
In contrast with this data, it has also been reported that subjects with the MC4R rs17782313 SNP exhibited a positive association with BMI (p = 0.018).2 Numerous studies reported that MC4R variants are associated with the incidence of obesity.35–37
FTO gene expressed at high levels in brain and hypothalamus, a region known to be responsible for appetite regulation.38 Biochemical studies have shown that the mutation in the FTO gene as an obesity susceptibility gene may lead to an increased risk of obesity.14,15,27,28,30,39 A recent report suggested that the obesity-associated elements within FTO region interact with an iroquois-class homeodomain protein 3 (IRX3) gene promoter and might be controlling expression of IRX3, therefore, it is possible that the IRX3 gene is also associated to obesity.40 In addition, mutations to obesity-related FTO introns of the FTO gene are associated with increased risk of many chronic diseases, including type II diabetes and cardiovascular disease.26,39,41 The IRX3 promoter, a gene several hundred thousand base pairs away, interacts with obesity-related FTO introns as well as a large number of other elements, therefore these introns act as regulatory elements of IRX3 expression, although evidence shows that the FTO gene itself does not play a role this interaction.40 Mutations within regulatory elements, spanning from chromosomal abnormalities include translocations, deletions, duplications, inversions, and aneuploidies to SNP cause diverse human diseases.42
MC4R is an important obesity candidate gene. This receptor consisting of 322-amino acids is encoded by a single exon located on chromosome 18q22 and is expressed at highest levels in the brain.43 Mutations in the MC4R gene contribute to disruption in energy homeostasis, weight gain, and the development of obesity.44 Furthermore, MC4R in the central nervous system plays a key role in regulation of glucose homeostasis. Thus, mutations in the human MC4R gene could affect the level of insulin secretion and leads to hyperinsulinemia.45 Extensive studies of the MC4R gene polymorphism showed that, among numerous variants, the rs17782313 genotype was significantly more frequent and associated with obesity.20,31,45–47 The mechanism of the association for rs17782313 polymorphism with BMI has not yet been elucidated and requires further study. Nevertheless, various studies suggest that the correlation between MC4R rs17782313 variant and high energy intake is significant.24,44,46
Conclusions
Obesity has a multifactorial origin and its prevalence has increased dramatically, thus determining the genetic polymorphism of genes might be an important approach as a marker of genetic predisposition to obesity for all population groups. Our data confirms previous observations that A-A genotype carriers have a higher risk for obesity compared with A-T and T-T genotype carriers. This knowledge may have important implications in personalized lifestyle management strategies to prevent obesity in genetically susceptible individuals. Finally, these results suggest that evaluation of FTO and MC4R genetic polymorphisms could be considered a prognostic tool to identify people at higher risk of developing obesity.
Disclosure
All authors declared no conflicts of interest. No funding was received for this study.
Acknowledgements
The authors would like to thank Iranian National Science Foundation and the National Institute of Genetic Engineering and Biotechnology, Tehran, Iran. The authors express their appreciation to all of the participants for donating their blood samples.
references
- 1. World Health Organization. Obesity: preventing and managing the global epidemic. Report of a WHO Consultation (WHO Technical Report Series 894); 2000.
- 2. Kirac D, Kasimay Cakir O, Avcilar T, Deyneli O, Kurtel H, Yazici D, et al. Effects of MC4R, FTO, and NMB gene variants to obesity, physical activity, and eating behavior phenotypes. IUBMB Life 2016 Oct;68(10):806-816.
- 3. Prakash J, Mittal B, Srivastava A, Awasthi S, Srivastava N. Association of FTO rs9939609 SNP with obesity and obesity-associated phenotypes in a north Indian population. Oman Med J 2016 Mar;31(2):99-106.
- 4. Cefalu WT, Bray GA, Home PD, Garvey WT, Klein S, Pi-Sunyer FX, et al. Advances in the science, treatment, and prevention of the disease of obesity: reflections from a diabetes care Editors’ expert forum. Diabetes Care 2015 Aug;38(8):1567-1582.
- 5. Simonds SE, Cowley MA, Enriori PJ. Leptin increasing sympathetic nerve outflow in obesity: A cure for obesity or a potential contributor to metabolic syndrome? Adipocyte 2012 Jul;1(3):177-181.
- 6. Jayawardena R, Byrne NM, Soares MJ, Katulanda P, Yadav B, Hills AP. High dietary diversity is associated with obesity in Sri Lankan adults: an evaluation of three dietary scores. BMC Public Health 2013 Apr;13(1):314.
- 7. Azadbakht L, Esmaillzadeh A. Dietary diversity score is related to obesity and abdominal adiposity among Iranian female youth. Public Health Nutr 2011 Jan;14(1):62-69.
- 8. Jafari-Adli S, Jouyandeh Z, Qorbani M, Soroush A, Larijani B, Hasani-Ranjbar S. Prevalence of obesity and overweight in adults and children in Iran; a systematic review. J Diabetes Metab Disord 2014 Dec;13(1):121.
- 9. Manna P, Jain SK. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: causes and therapeutic strategies. Metab Syndr Relat Disord 2015 Dec;13(10):423-444.
- 10. Zhao J, Bradfield JP, Li M, Wang K, Zhang H, Kim CE, et al. The role of obesity-associated loci identified in genome-wide association studies in the determination of pediatric BMI. Obesity (Silver Spring) 2009 Dec;17(12):2254-2257.
- 11. den Hoed M, Ekelund U, Brage S, Grontved A, Zhao JH, Sharp SJ, et al. Genetic susceptibility to obesity and related traits in childhood and adolescence: influence of loci identified by genome-wide association studies. Diabetes 2010 Nov;59(11):2980-2988.
- 12. Park S, Daily JW, Zhang X, Jin HS, Lee HJ, Lee YH. Interactions with the MC4R rs17782313 variant, mental stress and energy intake and the risk of obesity in Genome Epidemiology Study. Nutr Metab (Lond) 2016 May;13(1):38.
- 13. Speliotes EK, Willer CJ, Berndt SI, Monda KL, Thorleifsson G, Jackson AU, et al; MAGIC; Procardis Consortium. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet 2010 Nov;42(11):937-948.
- 14. Wu Q, Saunders RA, Szkudlarek-Mikho M, Serna IdeL, Chin K-V. The obesity-associated Fto gene is a transcriptional coactivator. Biochem Biophys Res Commun 2010 Oct;401(3):390-395.
- 15. Jia G, Yang C-G, Yang S, Jian X, Yi C, Zhou Z, et al. Oxidative demethylation of 3-methylthymine and 3-methyluracil in single-stranded DNA and RNA by mouse and human FTO. FEBS Lett 2008 Oct;582(23-24):3313-3319.
- 16. Church C, Lee S, Bagg EA, McTaggart JS, Deacon R, Gerken T, et al. A mouse model for the metabolic effects of the human fat mass and obesity associated FTO gene. PLoS Genet 2009 Aug;5(8):e1000599.
- 17. Fischer J, Koch L, Emmerling C, Vierkotten J, Peters T, Brüning JC, et al. Inactivation of the Fto gene protects from obesity. Nature 2009 Apr;458(7240):894-898.
- 18. McMurray F, Church CD, Larder R, Nicholson G, Wells S, Teboul L, et al. Adult onset global loss of the fto gene alters body composition and metabolism in the mouse. PLoS Genet 2013;9(1):e1003166.
- 19. Church C, Moir L, McMurray F, Girard C, Banks GT, Teboul L, et al. Overexpression of Fto leads to increased food intake and results in obesity. Nat Genet 2010 Dec;42(12):1086-1092.
- 20. Yilmaz Z, Davis C, Loxton NJ, Kaplan AS, Levitan RD, Carter JC, et al. Association between MC4R rs17782313 polymorphism and overeating behaviors. Int J Obes (Lond) 2015 Jan;39(1):114-120.
- 21. Loos RJ, Lindgren CM, Li S, Wheeler E, Zhao JH, Prokopenko I, et al; Prostate, Lung, Colorectal, and Ovarian (PLCO) Cancer Screening Trial; KORA; Nurses’ Health Study; Diabetes Genetics Initiative; SardiNIA Study; Wellcome Trust Case Control Consortium; FUSION. Common variants near MC4R are associated with fat mass, weight and risk of obesity. Nat Genet 2008 Jun;40(6):768-775.
- 22. Kochetova OV, Korytina GF, Akhmadishina LZ, Semenov EE, Viktorova TV. [Association of polymorphic variants of FTO and MC4R genes with obesity in a Tatar population]. Genetika 2015 Feb;51(2):248-255.
- 23. Horstmann A, Kovacs P, Kabisch S, Boettcher Y, Schloegl H, Tönjes A, et al. Common genetic variation near MC4R has a sex-specific impact on human brain structure and eating behavior. PLoS One 2013 Sep;8(9):e74362.
- 24. Khalilitehrani A, Qorbani M, Hosseini S, Pishva H. The association of MC4R rs17782313 polymorphism with dietary intake in Iranian adults. Gene 2015 Jun;563(2):125-129.
- 25. Qi L, Kraft P, Hunter DJ, Hu FB. The common obesity variant near MC4R gene is associated with higher intakes of total energy and dietary fat, weight change and diabetes risk in women. Hum Mol Genet 2008 Nov;17(22):3502-3508.
- 26. Liu G, Zhu H, Lagou V, Gutin B, Stallmann-Jorgensen IS, Treiber FA, et al. FTO variant rs9939609 is associated with body mass index and waist circumference, but not with energy intake or physical activity in European- and African-American youth. BMC Med Genet 2010 Apr;11(1):57.
- 27. Almén MS, Jacobsson JA, Moschonis G, Benedict C, Chrousos GP, Fredriksson R, et al. Genome wide analysis reveals association of a FTO gene variant with epigenetic changes. Genomics 2012 Mar;99(3):132-137.
- 28. Xi B, Shen Y, Zhang M, Liu X, Zhao X, Wu L, et al. The common rs9939609 variant of the fat mass and obesity-associated gene is associated with obesity risk in children and adolescents of Beijing, China. BMC Med Genet 2010 Jul;11(1):107.
- 29. Berulava T, Horsthemke B. The obesity-associated SNPs in intron 1 of the FTO gene affect primary transcript levels. Eur J Hum Genet 2010 Sep;18(9):1054-1056.
- 30. Gerken T, Girard CA, Tung Y-C, Webby CJ, Saudek V, Hewitson KS, et al. The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science 2007 Nov;318(5855):1469-1472.
- 31. Bjørnland T, Langaas M, Grill V, Mostad IL. Assessing gene-environment interaction effects of FTO, MC4R and lifestyle factors on obesity using an extreme phenotype sampling design: Results from the HUNT study. PLoS One 2017 Apr;12(4):e0175071.
- 32. Duicu C, Mărginean CO, Voidăzan S, Tripon F, Bănescu C. FTO rs 9939609 SNP is associated with adiponectin and leptin levels and the risk of obesity in a cohort of romanian children population. Medicine (Baltimore) 2016 May;95(20):e3709.
- 33. Illangasekera YA, Kumarasiri RP, Fernando DJ, Dalton CF. Association of FTO and near MC4R variants with obesity measures in urban and rural dwelling Sri Lankans. Obes Res Clin Pract 2016 Sep;10(Suppl 1):S117-S124.
- 34. Mangge H, Renner W, Almer G, Weghuber D, Möller R, Horejsi R. Rs9939609 variant of the fat mass and obesity-associated gene and trunk obesity in adolescents. J Obes 2011;2011:186368.
- 35. Xi B, Chandak GR, Shen Y, Wang Q, Zhou D. Association between common polymorphism near the MC4R gene and obesity risk: a systematic review and meta-analysis. PLoS One 2012;7(9):e45731.
- 36. McCaffery JM, Papandonatos GD, Peter I, Huggins GS, Raynor HA, Delahanty LM, et al; Genetic Subgroup of Look AHEAD; Look AHEAD Research Group. Obesity susceptibility loci and dietary intake in the Look AHEAD Trial. Am J Clin Nutr 2012 Jun;95(6):1477-1486.
- 37. Scherag A, Dina C, Hinney A, Vatin V, Scherag S, Vogel CI, et al. Two new Loci for body-weight regulation identified in a joint analysis of genome-wide association studies for early-onset extreme obesity in French and german study groups. PLoS Genet 2010 Apr;6(4):e1000916.
- 38. Labayen I, Ruiz JR, Huybrechts I, Ortega FB, Arenaza L, González-Gross M, et al. Dietary fat intake modifies the influence of the FTO rs9939609 polymorphism on adiposity in adolescents: The HELENA cross-sectional study. Nutr Metab Cardiovasc Dis 2016 Oct;26(10):937-943.
- 39. Martins MC, Trujillo J, Farias DR, Struchiner CJ, Kac G. Association of the FTO (rs9939609) and MC4R (rs17782313) gene polymorphisms with maternal body weight during pregnancy. Nutrition 2016 Nov-Dec;32(11-12):1223-1230.
- 40. Smemo S, Tena JJ, Kim K-H, Gamazon ER, Sakabe NJ, Gómez-Marín C, et al. Obesity-associated variants within FTO form long-range functional connections with IRX3. Nature 2014 Mar;507(7492):371-375.
- 41. Hsiao T-J, Lin E. Association of a common rs9939609 variant in the fat mass and obesity-associated (FTO) gene with obesity and metabolic phenotypes in a Taiwanese population: a replication study. J Genet 2016 Sep;95(3):595-601.
- 42. Chatterjee S, Ahituv N. Gene regulatory elements, major drivers of human disease. Annu Rev Genomics Hum Genet 2017 Aug;18:45-63.
- 43. Razquin C, Marti A, Martinez JA. Evidences on three relevant obesogenes: MC4R, FTO and PPARγ. Approaches for personalized nutrition. Mol Nutr Food Res 2011 Jan;55(1):136-149.
- 44. Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, et al. Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell 2005 Nov;123(3):493-505.
- 45. Rossi J, Balthasar N, Olson D, Scott M, Berglund E, Lee CE, et al. Melanocortin-4 receptors expressed by cholinergic neurons regulate energy balance and glucose homeostasis. Cell Metab 2011 Feb;13(2):195-204.
- 46. Corella D, Ortega-Azorín C, Sorlí JV, Covas MI, Carrasco P, Salas-Salvadó J, et al. Statistical and biological gene-lifestyle interactions of MC4R and FTO with diet and physical activity on obesity: new effects on alcohol consumption. PLoS One 2012;7(12):e52344.
- 47. Srinivasan S, Lubrano-Berthelier C, Govaerts C, Picard F, Santiago P, Conklin BR, et al. Constitutive activity of the melanocortin-4 receptor is maintained by its N-terminal domain and plays a role in energy homeostasis in humans. J Clin Invest 2004 Oct;114(8):1158-1164.